
Holoepipelágico (organismo)..............................................................................................................................................................Holoepipelagic
Holoépipélagique (organisme) / Holoepipelágico (organismo) / Holoepipelagic (immer in der epipelagic Zone) / holoepipelagic(总是,在epipelagic区) / Голоэпипелагический (организм) / Holoepipelagic (sempre nella zona epipelagica) /
Organismo, em geral um peixe, que habita de maneira, permanente, na zona epipelágica.
Ver: « Pelágico (organismo) »
&
« Epipelágica (zona) »
&
« Epifauna »
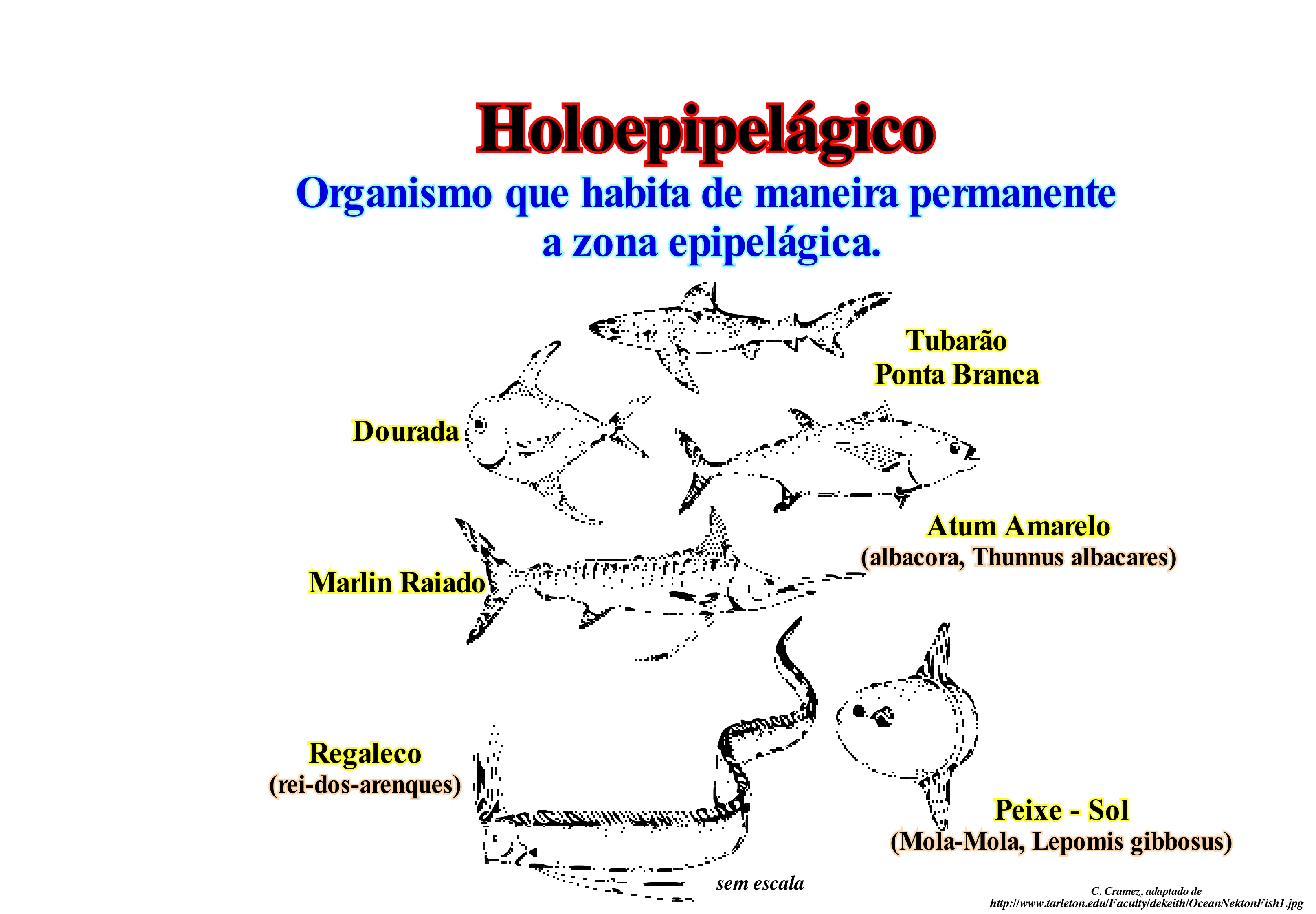
O nécton é constituído por bons nadadores, isto é, por organismos que não dependem das correntes marinhas para se deslocar. O nécton é, predominantemente, formado por animais vertebrados. Os peixes são de longe os mais numerosos, mas cada classe de vertebrados, excepto os anfíbios estão representados no nécton. O nécton oceânico está distribuído na zona de águas, relativamente, rasas. O nécton oceânico é composta por uma grande variedade de peixes ósseos, tubarões, raias, e, em menor medida, mamíferos e répteis. Os únicos invertebrados que podem ser considerados como nécton são os moluscos cefalópodes. Os peixes que passam as suas vidas nos ambientes epipelágicos são denominados holoepipelágicos. Entre eles, como ilustrados nesta figura, podem citar-se, por exemplo: os tubarões ponta branca, o besugo, o atum de barbatana amarela, o espadão estriado, o peixe agulha, o peixe-sol, etc. O segundo grupo de peixes oceânicos é chamado meroepipelágico. Estes peixes passam apenas parte de sua vida na zona epipelágica. Este é um grupo mais diversificado e inclui os peixes que passam suas vidas adultas nos ambientes epipelágicos, mas que desovam em águas costeiras (como o arenque, o tubarão-baleia, golfinho, etc.) ou em água doce (salmão). Um outro componente do nécton oceânico são os mamíferos marinhos. Entre eles estão as baleias (ordem dos cetáceos), as focas e leões marinhos (ordem Carnivora). Outros mamíferos marinhos incluindo o peixe-boi e dugongos (ordem Sirenia) e as lontras do mar (ordem Carnivora), mas estes ocupam as águas costeiras. Os répteis nectónicos oceânicos são quase, exclusivamente, as tartarugas e serpentes. Há também iguanas marinhos nas Ilhas Galápagos e crocodilos marinhos à volta de algumas ilhas na Indonésia e Austrália, mas estes répteis são litorais e animais costeiros. Muitos dos organismos que formam o nécton oceânico estão, perfeitamente, adaptados ao meio aquático. Outros meios como a redução dos ossos e a presença de uma camada de lípidios (gorduras e óleos) são utilizados pelos mamíferos marinhos para aumentar a flutuabilidade. Grandes quantidades de lípidios também estão presentes nos peixes nectónicos que não têm bexigas natatórias (tubarões, cavalas, bonitos). Além destes meios de produzir flutuabilidade, alguns animais nectónicos têm mecanismos hidrodinâmicos para a produção de flutuabilidade durante o movimento.
Holoplâncton.......................................................................................................................................................................................................................Holoplankton
Holoplancton / Holoplancton / Holoplankton / 终生浮游生物 / Голопланктон / Holoplankton /
Organismos que são plantónicos toda a sua vida, como, as diatomácias, radiolários, dinoflagelados, etc.
Ver: « Plâncton »
&
« Bentos »
&
« Megaplâncton »
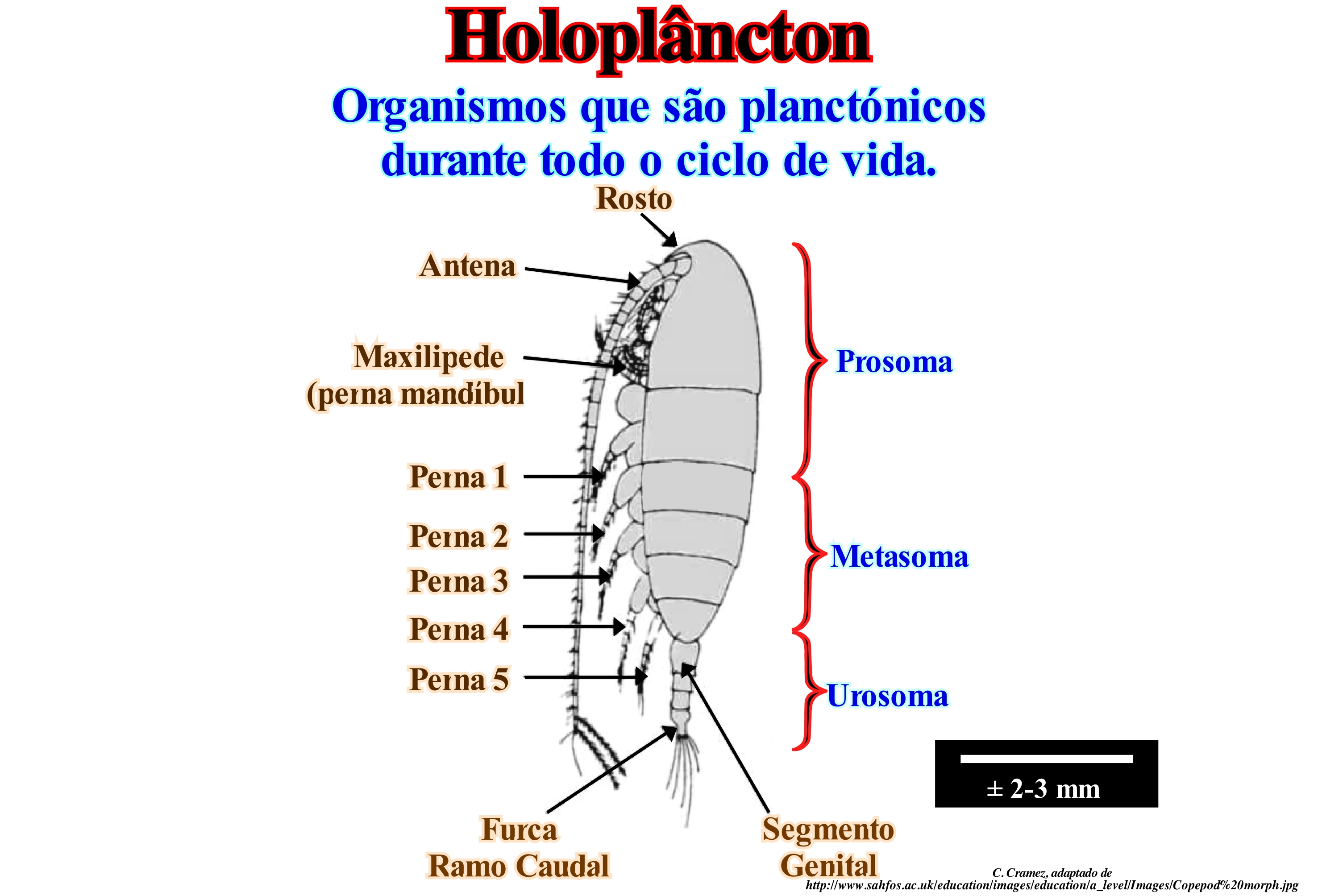
O holoplâncton é uma parte do plâncton animal ou zooplâncton. Chama-se holoplâncton ou plâncton verdadeiro aos corpos que permanecem no estado de plâncton ao longo do seu ciclo de vida. Ele é contrasta com o meroplâncton ou plâncton temporário, que é composto de organismos que passam apenas parte de suas vidas, habitualmente o estado larval, sob a forma de plâncton (medusas, crustáceos, larvas, vermes, etc.). O holoplâncton consiste, principalmente, de microalgas, animais unicelulares e alguns crustáceos como os copépodes, que representam cerca de 70% do zooplâncton. Os copépodos, como ilustrado nesta figura, são organismos planctónicos vivos livres (semelhante a uma pequena cochinilha, variedade de pulgões) que variam entre 1 e 10 milímetros de comprimento. Eles são capazes de circular livremente, sob pequenas distâncias, sacudindo os membros articulados e as antenas. Isto é interpretado como sendo um mecanismo para evitar predadores. As suas antenas ajudam, também, a evitar que eles se afundem através da coluna de água. Existem milhares de espécies de copépodos que formam o holoplâncton. Algumas vivem em água doce, mas a maioria são organismos marinhos, que têm uma morfologia básica. Os copépodos são, provavelmente, os organismos multicelulares mais abundantes na Terra. Certos autores pensam que existem cerca de 11500 espécies de copépodos divididos em aproximadamente 200 famílias e 1650 géneros. Os copépodos são um grupo diverso e, embora alguns sejam carnívoros ou onívoros, a maioria são herbívoros alimentando-se de fitoplâncton. Eles alimentam-se por meio de um sistema de filtragem ou utilizando os apêndices para manipular a célula. A emergência de populações de copépodos durante todo o ano afecta os ciclos de fitoplâncton no oceano. No holoplâncton entram-se, igualmente, cnidários (animais aquáticos conhecidos como celenterados, como as hidras de água doce, medusas, alforrecas, corais, anémonas-do-mar e as caravelas), quetognatas (pequenos animais marinhos com cerca de 15 cm de comprimento, de corpo alongado e não segmentado, com barbatanas laterais e caudais que são extensões da ectoderme), ostracodos (pequenos crustáceos dotados de uma carapaça calcária, composta por duas valvas que se articulam-se através de uma estrutura denominada charneira, localizada na região dorsal do animal), etc.
Homeostasia..............................................................................................................................................................................................................................Homeostasis
Homèostasie / Homeostasia / Homöostase / 稳态 /Гомеоста́з / Omeostasi /
Propriedade que um organismo tem de regular, fisiologicamente, o seu ambiente interno afim de garantir a sua estabilidade em resposta às flutuações do ambiente externo e clima.
Ver: « Anaeróbico (ambiente) »
&
« Teoria dos sistemas »
&
« Gaia (hipótese) »
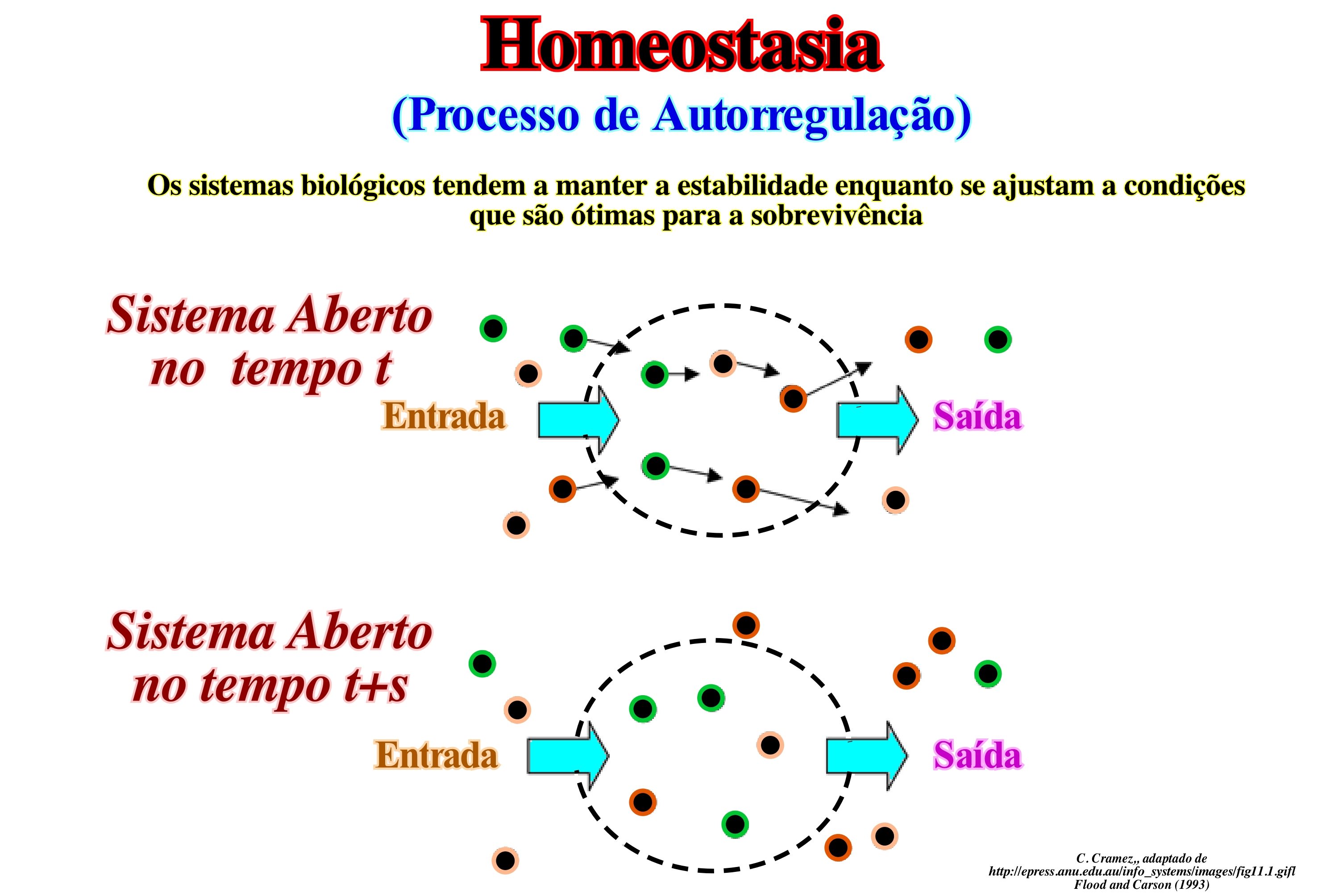
Homeostasia não é apenas uma das propriedades mais importantes de qualquer organismo vivo, mas também é aplicável a organizações humanas ou de trabalho tratados como sistemas abertos. Uma organização necessita de recrutar novos funcionários para substituir aqueles que se aposentam, mas também tem necessidade de matérias primas, energia e informações para uso dos seus processos e operações afim de manter um estado estável. Uma organização que aparece, externamente, estática e imutável, para os observadores externos, está, internamente, num estado de fluxos de equilíbrio dinâmico. Outro aspecto importante de um sistema aberto num estado de equilíbrio dinâmico é que ele depende de mecanismos de retroacção para permanecer nesse estado. Na hierarquia de Boulding, que classifica os sistemas de acordo com sua complexidade, não é surpreendente descobrir que propriedades exibidas pelos sistemas inferiores da hierarquia também são encontrados nos mais elevados, porque estes são construídos sobre os primeiros. Um sistema que é classificado como um sistema aberto, que possui todas as qualidades que pertencem ao sistema ao nível cibernético (autorregulação). O comportamento dos sistemas abertos é, em grande medida, determinado pelos mecanismos de retroacção neles presente. Existem dois tipos de retroacção que operam na maioria dos sistemas, ou seja, uma retroacção negativa e positiva. A primeira reduz ou elimina o desvio do sistema a partir de uma determinada norma. Um mecanismo de retroacção negativo tende a neutralizar o efeito de perturbação do ambiente para que o sistema possa manter o seu rumo normal de funcionamento. Por outro lado, ele amplifica a retroacção positiva ou acentua a mudança, o que leva a uma divergência contínua a partir do estado inicial. A retroacção positiva colabora com a retroacção negativa em sistemas vivos (por exemplo, organismos e organizações, também, ambos os tipos de retroacção estão presentes durante o crescimento, embora o resultado seja positivo). No entanto, a operação de retroacção positiva por si só pode, eventualmente, resultar num sistema de desintegração ou colapso. Uma retroacção negativa tem um papel fundamental na capacidade do sistema para atingir um estado de equilíbrio, ou homeostasia.
Homotaxia................................................................................................................................................................................................................................................Homotaxy
Homotaxie / Homotaxia / Homotaxy (Ähnlichkeit der Anordnung) / Homotaxy (相似的安排) / Гомотаксис / Homotaxy (somiglianza di sistemazione) /
Similaridade de um arranjo das partes de um todo. Analogia entre intervalos estratigráficos ou sucessões de fósseis em regiões diferentes. Analogia do arranjo dos estratos caracterizados por fósseis semelhantes que ocupam diferentes posições e sem conotação de idades. Analogia de arranjo de fósseis com a mesma posição relativa, mas que não são, necessariamente, contemporâneos. Termo introduzido para evitar a confusão entre uma analogia taxonómica e sincronismo.
Ver: « Fácies »
&
« Fóssil Característico »
&
« Idade Relativa »
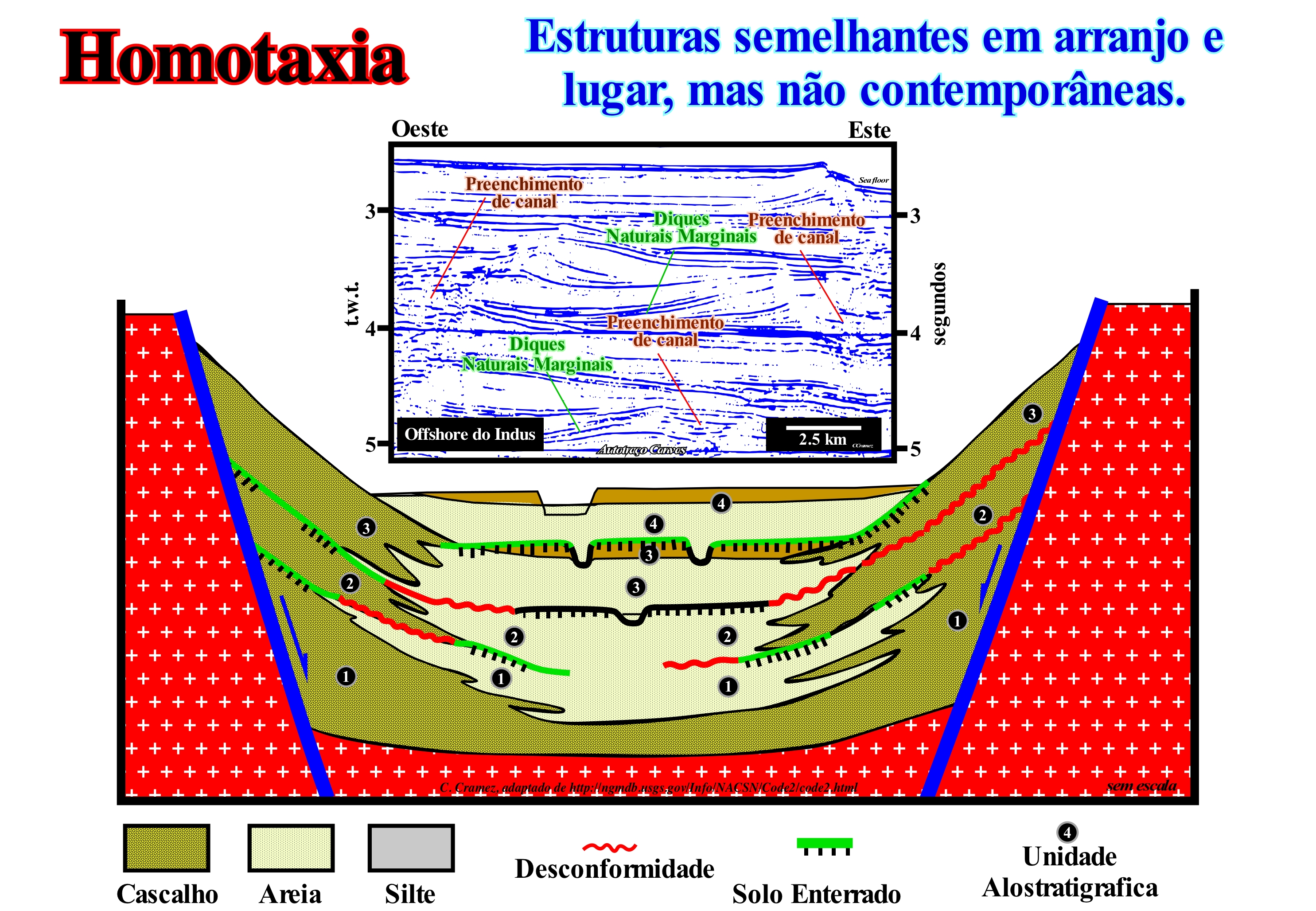
Homotaxia foi, inicialmente, proposto por Huxley (1982) para evitar a falácia de confundir uma semelhança taxonómica (com os mesmo grupos de organismos biológicos) com sincronismo. Assim, actualmente a homotaxia é muitas vezes definida como a similaridade taxonómica entre sequências estratigráficas ou fósseis em regiões separadas (Bates & Jackson, 1987), ou o facto que de estratos caracterizados por fósseis ocupem semelhantes posições em diferentes sequências verticais sem no entanto terem, necessariamente, a mesma idade. O termo homotaxia pode, também, ser utilizado par exprimir o facto que estratos ou camadas não contemporâneas (muitas vezes porque são muito afastadas uma das outras) possam conter os mesmos fósseis. Demonstrar que uma sucessão de fauna, flora ou taxa (plural de táxon) se pode confirmar de um corte geológico para outro, de uma bacia sedimentar para outra ou de um continente a outro, não é fácil e constitui o que, normalmente, muitos geocientistas chamam homotaxia. Observar os mesmo fósseis em dois afloramentos de bacias sedimentares diferentes não mostra, de maneira nenhuma, que esses afloramentos são síncronos. Na realidade, isto é, o grande problema dos fósseis guias (com uma grande distribuição espacial e que ocorrem durante um breve espaço de tempo) e a questão principal é de saber como é que se passa de eventos geológicos ordenados para a eventos geológicos datados. Este problema constitui o grande impasse da biostratigrafia. Os dados paleobiológicos e estratigráficos característicos de um determinado evento geológico não permitem estabelecer o seu sincronismo. Sobre este assunto não se pode esquecer que: (i) Os fósseis significam uma sucessão de eventos irreversíveis e (ii) As correlações biostratigráficas dependem da emergência evolucionária de uma espécie, a partir do seu antepassado, da sua sobrevivência e expansão geográfica. O principio das correlações assume que as comunidades biológicas, em diferentes lugares, são sincronizadas ou que os sinais biológicos são transmitidos, instantaneamente, ou ambos, o que não é, necessariamente, verdade. Scott, G. H. (1985) considerou que o principal problema na biostratigrafia é a justificação do uso de fósseis para identificar horizontes isócronos e propôs uma primeira solução, uma vez que os geocientistas estabelecem sucessões de eventos fósseis, mas não há nenhuma justificação teórica para considerar que a mesma posição estratigráfica (homotaxia) corresponde a um mesmo tempo de deposição. Ele propôs uma primeira solução usando a homotaxia como teste de diacronismo, quer isto dizer, que os eventos que não mantém uma ordem estratigráfica invariante são considerados como diacrónicos. Os eventos que resistem a este teste não são considerados isócronos, mas o diacronismo potencial das suas distribuições individuais é, inversamente, relacionado com o seu espaçamento estratigráfico. Quanto mais perto estão os eventos homotaxiais no espaço estratigráfico, menos eles estão separados no tempo. Assim, um dos principais objectivos da investigação biostratigráfica deve ser o de fazer testes de homotaxia mais rigorosos aumentando a densidade de eventos. O esquema ilustrado nesta figura, que é um bom exemplo de uma interpretação alostratigráfica (unidades estratigráficas cartografáveis definidas e identificadas na base das descontinuidades que as limitam) dos depósitos aluviais e lacustres de uma bacia de tipo rifte, quer a bacia seja do Cretácico ou Neogénico, mostra que o conceito de homotaxia se pode aplicar, igualmente, às correlações litológicas, para designar o facto que estratos caracterizados por relações geométricas e posições semelhantes em diferentes sequências verticais independentemente da sua idade. De facto, neste corte geológico, os depósitos aluviais e lacustres podem ser incluídos numa única formação geológica, ou podem ser separados lateralmente em formações geológicas distintas na base dos contrastes de litologia (cascalho, argila). As mudanças de litologia são abruptas e acentuadas, tanto vertical como lateralmente. A homotaxia não implica, necessariamente, um sincronismo dos depósitos. Os depósitos aluviais e lacustres, por exemplo, podem ser incluídos numa única formação geológica ou ser separados, lateralmente, em formações distintas, na base das relações geométricas ou na base dos seus fósseis. Contudo, quatro unidades alostratigráficas, cada uma incluindo duas ou três fácies, podem ser definidas com base nas descontinuidades lateralmente cartografáveis. Como se pode constatar no autotraço de um detalhe de uma linha sísmica regional do Indus, é evidente a similaridade e o diacronismo dos cones submarinos de talude (estruturas em asas de gaivota de P. Vail) constituem, globalmente, um bom exemplo de homotaxia.
Horizonte (geológico, sísmico)...............................................................................................................................................................................................Horizon
Horizon (géologique, sismique) / Horizonte (geológico, sísmico) / Horizont (geologischen, seismischen) / 地平线(地质,地震) / Горизонт (геологический, сейсмический) / Orizzonte (geologico, sismico) /
Camada ou um conjunto de camadas com uma composição particular e fósseis característicos, que permitem de lhe dar uma idade relativa. Nas linhas sísmicas, um horizonte é um reflector, mais ou menos, contínuo, que sublinha uma interface entre dois intervalos estratigráficos com impedâncias acústicas diferentes (produto da velocidade das ondas sísmicas através um intervalo pela densidade do mesmo).
Ver: « Reflexão Sísmica »
&
« Secção Geológica »
&
« Camada de Referência »
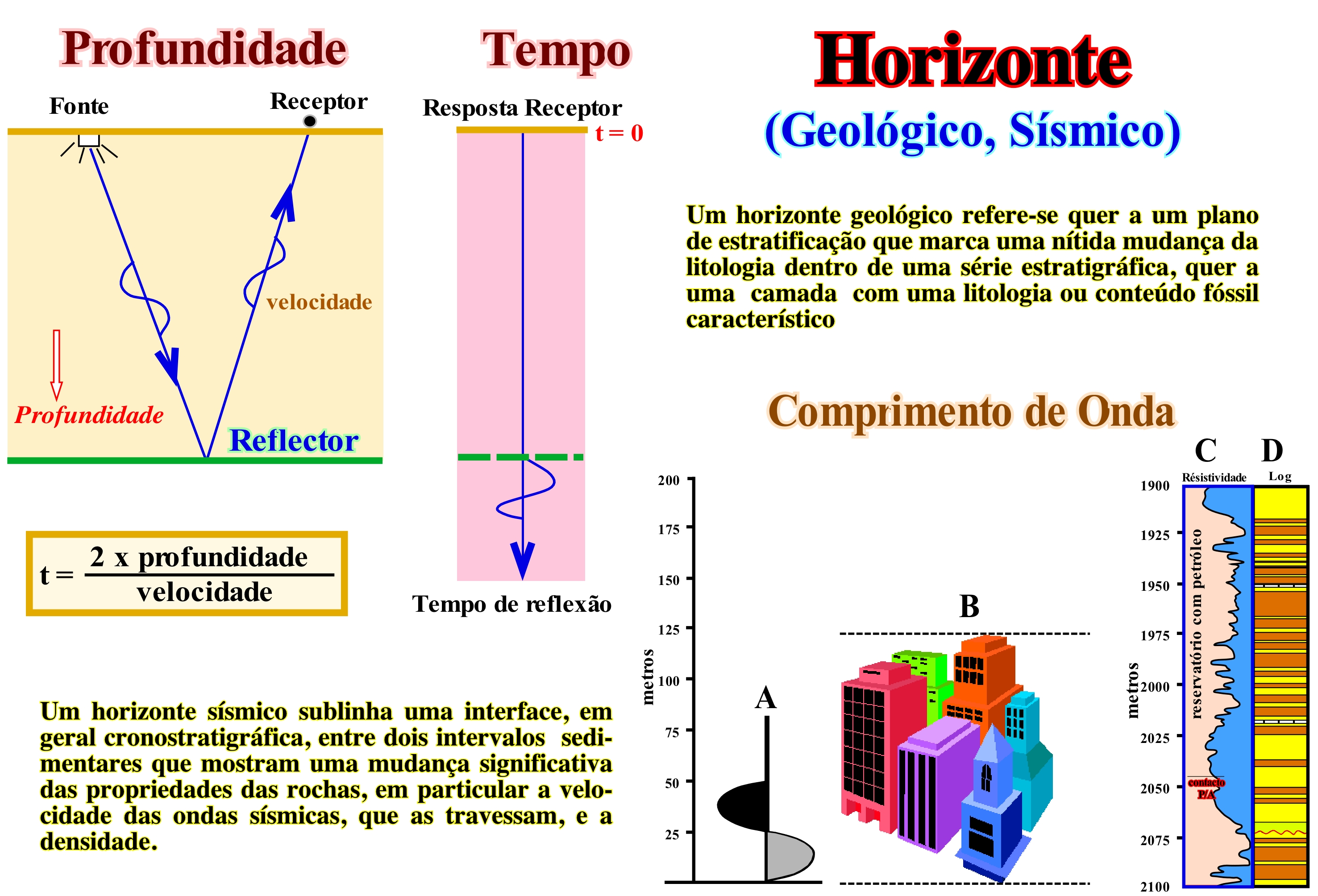
Estes esquemas ilustram, em profundidade e em tempo, um dos mecanismos básicos da sísmica de reflexão, o qual nunca deve ser esquecido quando os geocientistas interpretam as linhas sísmicas em termos geológicos. À esquerda, no esquema “Profundidade” pode ver-se a fonte de energia, a profundidade de um reflector (interface entre dois intervalos sedimentares), o receptor (geofone) e trajecto das ondas sísmicas, materializado pelo trajecto de um raio, enquanto que no esquema “Tempo” se encontra resposta em tempo. O tempo gasto por uma onda é igual a duas vezes a profundidade do reflector, ou seja, ida e volta (ida = fonte / reflector e volta = reflector / geofone) a dividir pela velocidade das ondas no meio considerado. A resposta em profundidade é dada pela profundidade do reflector em tempo multiplicada pela velocidade das ondas sísmicas dentro do intervalo sedimentar sobrejacente ao reflector. No esquema da direita “Comprimento de Onda”, está ilustrada uma comparação entre (A) um ciclo de uma onda sísmica de 30 Hz*, num meio de velocidade 1832 m/s (60 Hz com 3 500 m/s), (B) altura dos edifícios de uma cidade e (C) a coluna estratigráfica atravessada por um poço de pesquisa numa área onde foi encontrado uma acumulação económica de hidrocarbonetos, ou seja, um campo petrolífero. Um ciclo de uma onda sísmica normal é, quase sempre, inferior a um prédio de 50 m de altura. Quando um geocientista propõe, numa tentativa de interpretação geológica de uma linha sísmica, corpos geológicos de pequeno tamanho como, o preenchimento de um vale cavado, uma barra de meandro, etc., ele têm, imperativamente, que tomar em linha conta as escalas e a resolução das linhas sísmica. A resolução sísmica vertical, que corresponde à distância vertical mínima entre duas interfaces, necessária para que uma reflexão simples possa ser observada, raramente, é inferior a 50 m. Ela é controlada, principalmente, pelo comprimento de onda do sinal sísmico. À medida que a espessura de um intervalo se aproxima de um quarto de comprimento de onda, a interferência construtiva produz uma acumulação de amplitude conhecida como o efeito de sintonização, cuja espessura é, geralmente, considerada como o limite de resolução ou da separabilidade como dizem certos geocientistas. Como o comprimento de onda é igual à velocidade dividida pela frequência, um quarto de comprimento de onda representa cerca de 7 m para intervalos pouco profundos com uma velocidade de, mais ou menos, 1 828 m/s, para uma onduleta (“wavelet” dos geocientistas de língua inglesa, que é uma função capaz de decompor e descrever ou de representar outra função, originalmente, descrita em tempo, de forma a que s e possa analisar esta outra função em diferentes escalas de frequência e de tempo) de 60 Hz ou de, mais ou menos, 76 metros para intervalos profundos com uma velocidade de 4500 m/s e uma onduleta de 15 Hz. A resolução diminui com a profundidade porque a velocidade aumenta e frequência diminui. A resolução sísmica lateral é determinada pelo raio da zona de Fresnel a qual depende do comprimento de onda da onduleta (pulso acústico) e da profundidade do reflector. De facto, As ondas sísmicas que dão origem a um evento de reflexão são reflectidas a partir de uma área bastante grande, aproximadamente circular da interface reflectora conhecida como a primeira zona de Fresnel. Reflexões desta zona chegam a um geofone para interferir construtivamente. O raio desta zona é muitas vezes tomado como a resolução horizontal para dados sísmicos não migrados. Para uma onduleta de 60 Hz a 1706metros / segundo, este raio seria de cerca de 152 metros a uma profundidade de 1524 metros. O tamanho da zona de Fresnel, também, aumenta rapidamente com a profundidade. Como a migração colapsa zona de Fresnel, esta medida de resolução não é apropriada para os dados sísmicos migrados que são, actualmente, os mais usados para as tentativas de interpretação geológica das linhas sísmicas. Em princípio, depois da migração, a resolução horizontal é reduzida para traçar o espaçamento. Todavia, o ruído (mancha de migração de certos geocientistas), muitas vezes, torna-se um factor que ,pode reduzir de maneira significativa, a resolução. Outros factores podem contribuir para limitar a resolução lateral como, por exemplo, são a incerteza da velocidade de intervalo, a completude matemática do algoritmo de migração e amostragem espacial (intervalo entre os geofones).
(*) Unidade derivada de frequência do sistema internacional (SI), que é equivalente a um evento por segundo (s-1 ou 1 / s) e está ligado à velocidade (v) e o comprimento de onda (λ) pela relação: f = v/λ.
Humificação.............................................................................................................................................................................................................................Humification
Humification / Humificación / Humifizierung / 腐殖化 / Гумификация (образование гумуса) / Umificazione /
Degradação do material orgânico de um solo, o que causa, que alguns horizontes do solo se tornem castanhos ou mesmo negros.
Ver: « Solo »
&
« Humo »
&
« Matéria Orgânica (tipos) »
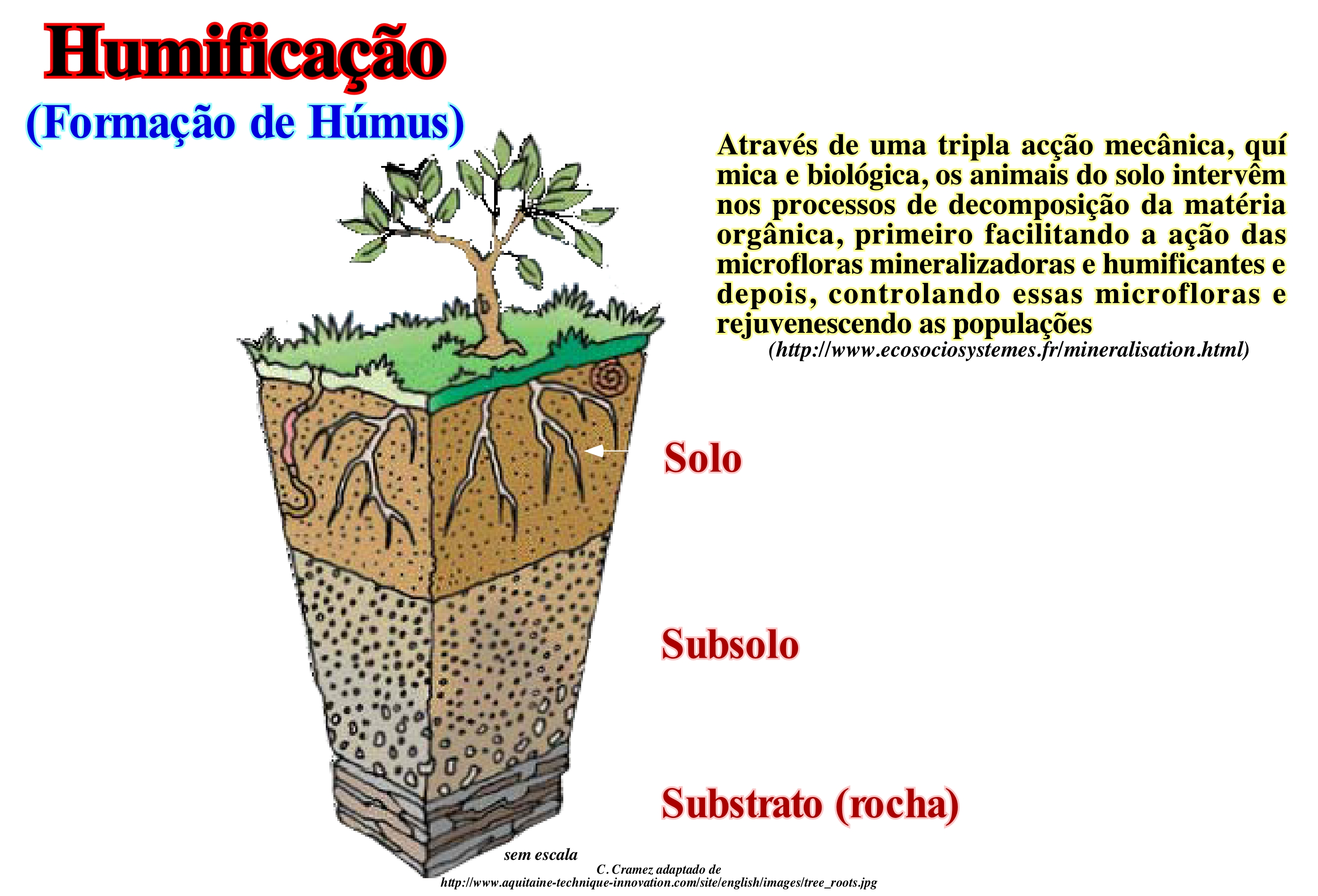
A humificação é o processo de transformação da matéria orgânica em húmus, sob a influência da microflora e microfauna. Ela sucede a decomposição de detritos orgânicos. Novas moléculas se desenvolvidos por via microbiana e físico-químicas (néossíntese). Esquematicamente, a formação de um solo pode dividir em três fases que se sobrepõem, mais ou menos, no tempo: (i) Alteração, isto é, a decomposição das rochas ; (ii) Humificação, ou seja, a introdução de matéria orgânica (húmus) e (iii) Diferenciação, que traduz as migrações e a acumulação de muitos elementos. A decomposição da rocha (rocha mãe do solo) fornece um material, mais ou menos móvel (pedologicamente a rocha-mãe), que é colonizado, gradualmente, por uma vegetação cada vez mais diferenciada. Isto traz de volta no terreno uma quantidade, mais ou menos, significativa, de resíduos, cuja transformação dá origem ao húmus. Alguns componentes do húmus, em parceria com os movimentos da água no solo, podem provocar deslocamentos das substâncias (migração), trazendo a aparência de camadas sobrepostas, algumas pobres e outras ricas em tais substâncias. Estas camadas são os horizontes, cujo conjunto constitui o perfil do solo. O horizonte A é o horizonte superficial com matéria orgânica (restos de plantas). O horizonte C corresponde à rocha pouco ou ligeiramente alterada e os horizontes B são os horizontes intermediários que aparecem nos solos evoluídos. Os solos são imaturos ou pouco evoluídos tem perfil de AC, enquanto que os solos evoluídos têm um perfil ABC. Os horizontes B são formados pelo intemperismo das rochas ou pelos movimentos de matéria a partir do horizonte A. Note que um solo é uma película de alteração que recobre uma rocha. Ele é formado por uma fracção mineral e matéria orgânica (húmus). Um solo forma-se a partir da rocha, mas depois ele evolui sob a influência de factores ambientais, principalmente o clima e a vegetação. Certos geocientistas pensam que no processo de humificação existem ritmos independentes da temperatura, humidade, luz, variações de pressão atmosférica, microflora e variações químicas sazonais. Eles pensam, que certos factores de origem eléctrica e indirectamente da actividade solar podem ser responsáveis de tais ritmos.
Humo..................................................................................................................................................................................................................................................................................Humus
Humus / Humus / Humus / 腐殖质 / Гумус, перегной / Humus /
Matéria orgânica de um solo que atingiu o ponto de estabilidade, a partir do qual ela não se pode decompor mais e fica, se as condições ambientais não mudarem, nesse estado durante séculos se não milénios.
Ver: « Solo »
&
« Humificação »
&
« Matéria Orgânica (tipos) »

Húmus ou humo é a matéria orgânica depositada no solo, resultante da decomposição de animais e plantas mortas, ou de seus subprodutos. O processo de formação do húmus é chamado humificação e pode ser natural, quando produzido espontâneamente por bactérias e fungos do solo (os organismos decompositores), ou artificial, quando o homem induz a produção de húmus, adicionando produtos químicos e água a um solo pouco produtivo. Vários agentes externos como a humidade e a temperatura contribuem para a humificação. Na formação do húmus há libertação de diversos nutrientes, mas é de especial consideração a libertação de nitrogénio. A compostagem é uma forma de "fabricar" húmus para utilizar como composto, ou seja, fertilizante orgânico na agricultura. A vermicompostagem é o uso da minhoca na produção de húmus, decompondo resíduos e dejectos de animais e também o lixo urbano (orgânico), colaborando com a melhoria dos solos, sequestrando carbono e eliminando cheiros desagradáveis. A vermicompostagem é um processo bastante difundido, em especial entre moradores de áreas rurais, visto a minhoca ser uma verdadeira máquina de limpeza dos resíduos. Quando colocada a quantidade correta de minhocas (ao redor de 5000 unidades por metro quadrado) em 30 a 35 dias (na compostagem normal leva de 100 a 300 dias), pode transformar 2,5 toneladas de resíduos orgânicos em húmus, em um canteiro de 10 x 0,80 x 0,40m. A minhoca come os resíduos, e seu excremento possui ao redor de 2 milhões de bactérias por grama, enriquecendo o solo deixando disponível as plantas, praticamente, todo o complexo mineral (cinco vezes e meia mais nitrogénio, duas vezes mais cálcio, duas vezes e meia mais magnésio, sete vezes mais fósforo e onze vezes mais potássio que o solo ou o resíduo que se alimentou). A formação do húmus é biogénica, ou seja, o húmus pode ser formado por oxidação simples de matéria orgânica morta, na ausência de organismos vivos, mas esse processo é bastante acelerado quando os organismos ingerem material orgânico ou secretam enzimas que a transformam.

