Professor Associado, Universidade Fernando Pessoa
| Outras vias metabólicas: | |||
| Química Orgânica: |
O piruvato produzido na glicólise ainda contém bastante poder redutor (verifique o estado de oxidação de cada um dos seus carbonos e compare-o com o estado de oxidação do carbono no CO2). Este poder redutor vai ser aproveitado pela célula no ciclo de Krebs. Em primeiro lugar, o piruvato é utilizado para produzir acetil-CoA, que é uma forma activada de acetato (CH3COO-)

Nesta reacção intervém a piruvato desidrogenase. É uma enzima bastante complexa, que contém bastantes cofactores: lipoamida, FAD, coenzima A. A hidrólise da ligação tioéster (S-C=O) do acetil-CoA é bastante exergónica, pelo que a sua formação exige energia. Essa energia provém da descarboxilação do piruvato (note que o piruvato tinha três carbonos e a porção acetil do acetilCoA apenas possui dois: o grupo carboxilato migrou como CO2). A energia proveniente de descarboxilações é frequentemente usada pela célula para empurrar um equilíbrio no sentido da formação de produtos, como se verá em várias reacções do ciclo de Krebs e na gluconeogénese.
Na primeira reacção do ciclo de Krebs, o acetil-CoA é adicionado a oxaloacetato, dando origem a citrato, numa reacção de adição aldólica. A hidrólise do tioéster ajuda a deslocar o equilíbrio no sentido da formação de produtos:

O citrato é depois isomerizado a isocitrato. Este é então descarboxilado a a-cetoglutarato. Se o citrato não tivesse sido isomerizado a isocitrato antes da descarboxilação, esta produziria um composto de carbono ramificado, mais difícil de metabolizar.

Tal como o piruvato, o a-cetoglutarato é um a-cetoácido, i.e., possui um grupo carbonilo adjacente ao grupo ácido carboxílico. É portanto de prever que reaja exactamente como o piruvato, i.e., que a sua descarboxilação forneça energia suficiente para que se forme uma ligação tioéster com a coenzima A. E é isto que de facto ocorre... A enzima responsável por esta reacção, a a-cetoglutarato desidrogenase, é aliás bastante análoga à piruvato desidrogenase na sua composição e cofactores.

A ligação tioéster do succinil-CoA é, como todas as ligações tioéster, bastante energética.A sua hidrólise vai constituir o único ponto do ciclo de Krebs onde ocorre produção directa de ATP (ou equivalente).
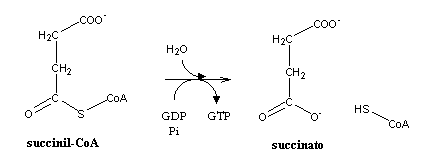
O succinato é, tal como o oxaloacetato, um produto com quatro carbonos. A parte final do ciclo de Krebs consiste em regenerar o oxaloacetato a partir do succinato. O succinato é primeiro oxidado a fumarato, pelo complexo succinato desidrogenase (também denominado complexo II), que se encontra na face matricial da membrana interna da mitocôndria. A oxidação de ligação simples a dupla (alcanos a alcenos) tem um potencial demasiado elevado para que os electrões possam ser aceites pelo NAD+ (E0=-320 mV). A célula utiliza portanto FAD (E0= 0 mV)como aceitador destes electrões. A hidratação do fumarato produz malato, que depois é oxidado a oxaloacetato, completando o ciclo. Uma sequência semelhante de reacções ocorre na b-oxidação dos lípidos.

O resultado do ciclo de Krebs é portanto:
Acetil-CoA + oxaloacetato + 3 NAD+ + GDP + Pi +FAD --> oxaloacetato + 2 CO2 + FADH2 + 3 NADH + 3 H+ + GTP
 |
Biochemistry,
by Donald Voet & Judith Voet Um excelente livro. Expõe a Bioquímica com referências constantes à química orgânica e à lógica bioquímica. A inspiração destas páginas.... Particularmente indicado a estudantes de Licenciaturas em Bioquímica, Ciências Farmacêuticas ou Química. |
 |
Biochemistry,
Stryer Um texto clássico, frequentemente actualizado e re-editado. |
 |
Textbook
of Biochemistry with Clinical Correlations, Thomas Devlin Aconselhado a estudantes de licenciaturas em Medicina, Enfermagem, etc. Imensos exemplos da aplicação da bioquímica a casos clínicos |
 |
Principles
of Biochemistry, Lehninger Um texto clássico, frequentemente actualizado e re-editado. |